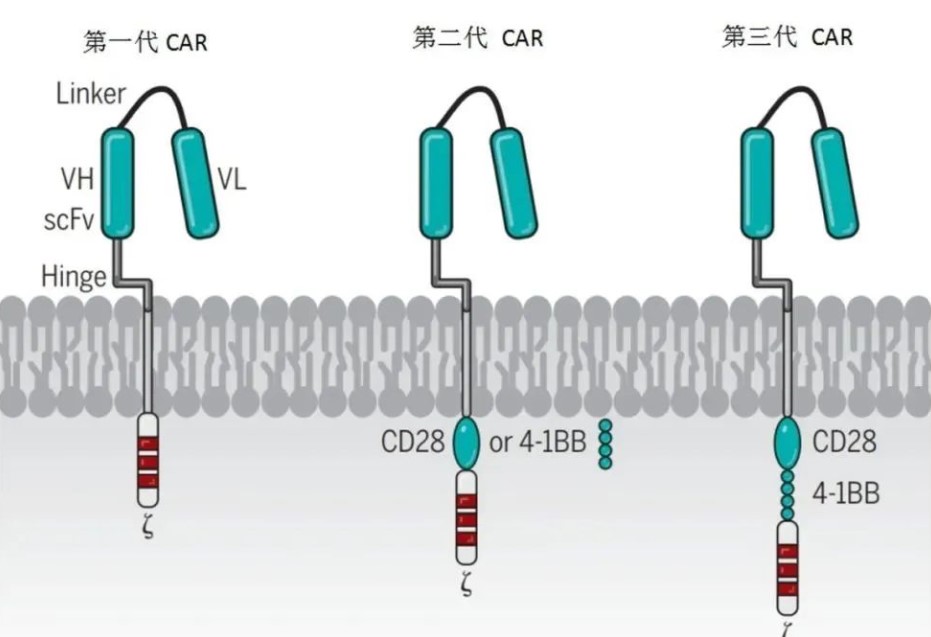
CAR-T免疫疗法
是一种过继性细胞免疫疗法,它是利用基因技术,在T细胞内导入一个可以将CAR表达于其表面的融合基因,并使T细胞发挥作用,克服肿瘤局部免疫抑制微环
境,打破宿主免疫耐受状态,靶向杀伤肿瘤细胞。
这篇文章小编将具体讲解一下CAR受体的结构的四个主要部分的功能。
PART 1
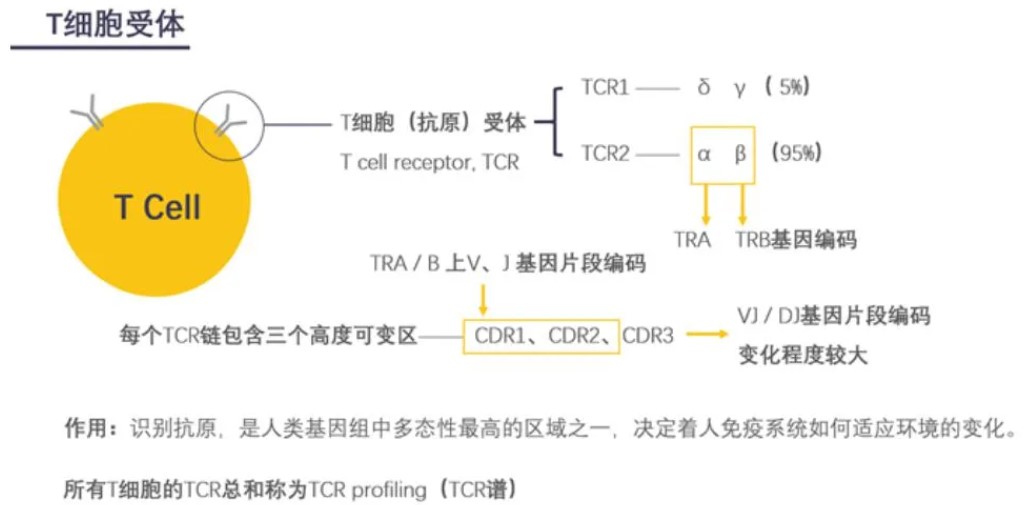
什么是TCR
T 细胞受体(T-cell receptor),简称 TCR,是在 T 细胞表面发现的一种分子[1],T 细胞介导的抗原识别依赖 于 T
细胞受体与抗原-主要组织相容性复合体(MHC)的相互作用(图 1)。当 TCR-CD3 复合物、抗原肽、MHC (HLA)结合时,T
淋巴细胞通过信号转导被激活,进入后续的免疫应答过程。

TCR 结构图
TCR为所有T细胞表面的特征性标志,以非共价键与CD3结合,形成TCR-CD3复合物。TCR的作用是识别抗原。TCR是由两条不同肽链构成的异二聚
体,由α、β两条肽链组成,每条肽链又可分为可变区(V区),恒定区(C区),跨膜区和胞质区等几部分;其特点是
胞质区很短。
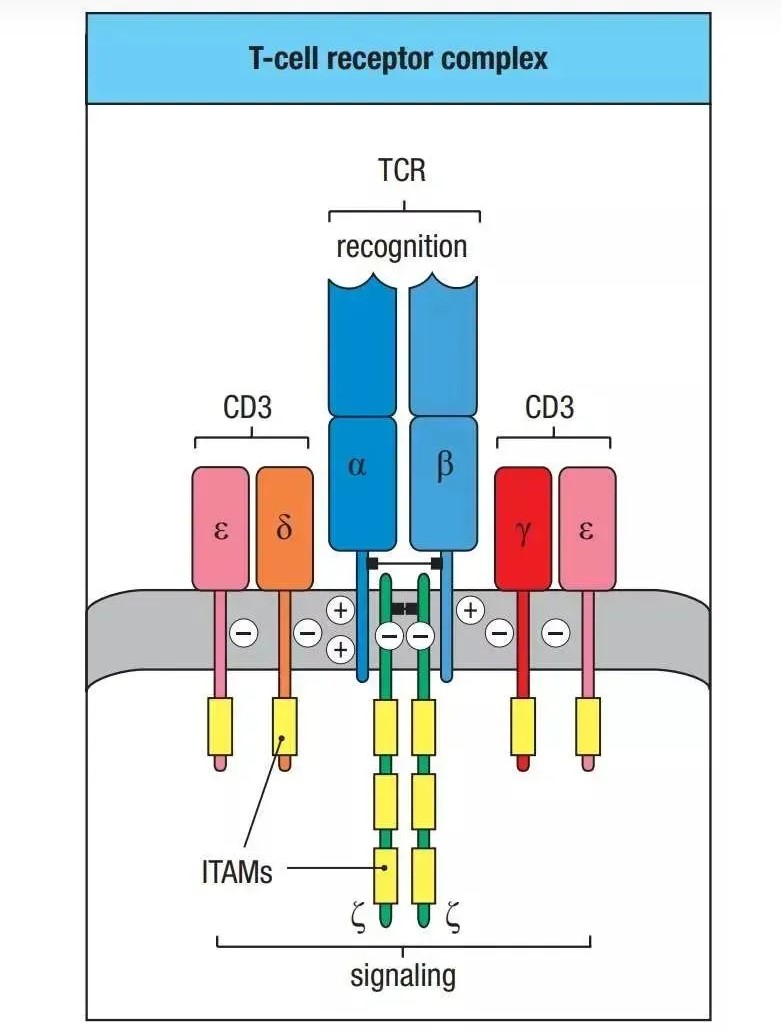
TCR-CD3结构
TCR分子属于免疫球蛋白超家族,其抗原特异性存在于V区;V区(Vα、Vβ)又各有三个高变区CDR1、
CDR2、CDR3,其中以CDR3变异最大,直接决定了TCR的抗原结合特异性。在TCR识别MHC-抗原肽复合体时,CDR1,CDR2识别和结合
MHC分子抗原结合槽的侧壁,而CDR3直接与抗原肽相结合。TCR分为两类:TCR1和TCR2;TCR1由γ和&
amp;
amp;
amp;
amp;
amp;
amp;
amp;
amp;
amp;
amp;
amp;
amp;
amp;
amp;
delta;两条链组成,TCR2由α和β两条链组成。外周血中,90%-95%的T细胞表达TCR2;而且任一
T细胞只表达TCR2和TCR1之一。
TCR 的α链和β链的 V(D)J 重组
TCR 是高度多样化的异源二聚体,人类约 95%的 T
细胞表达α链和β链(αβ-TCR,两条链分别由 TRA 和 TRB
编码),外周血 T 细胞(约占 1–5%)和粘膜位置 T
细胞还会表达γδ链(γδ-TCR,分别由 TRG 和 TRD
编码) ,在肿瘤发生和处于某种疾病状态下,αβ-TCR
和γδ-TCR 比例会发生变化。
α chain
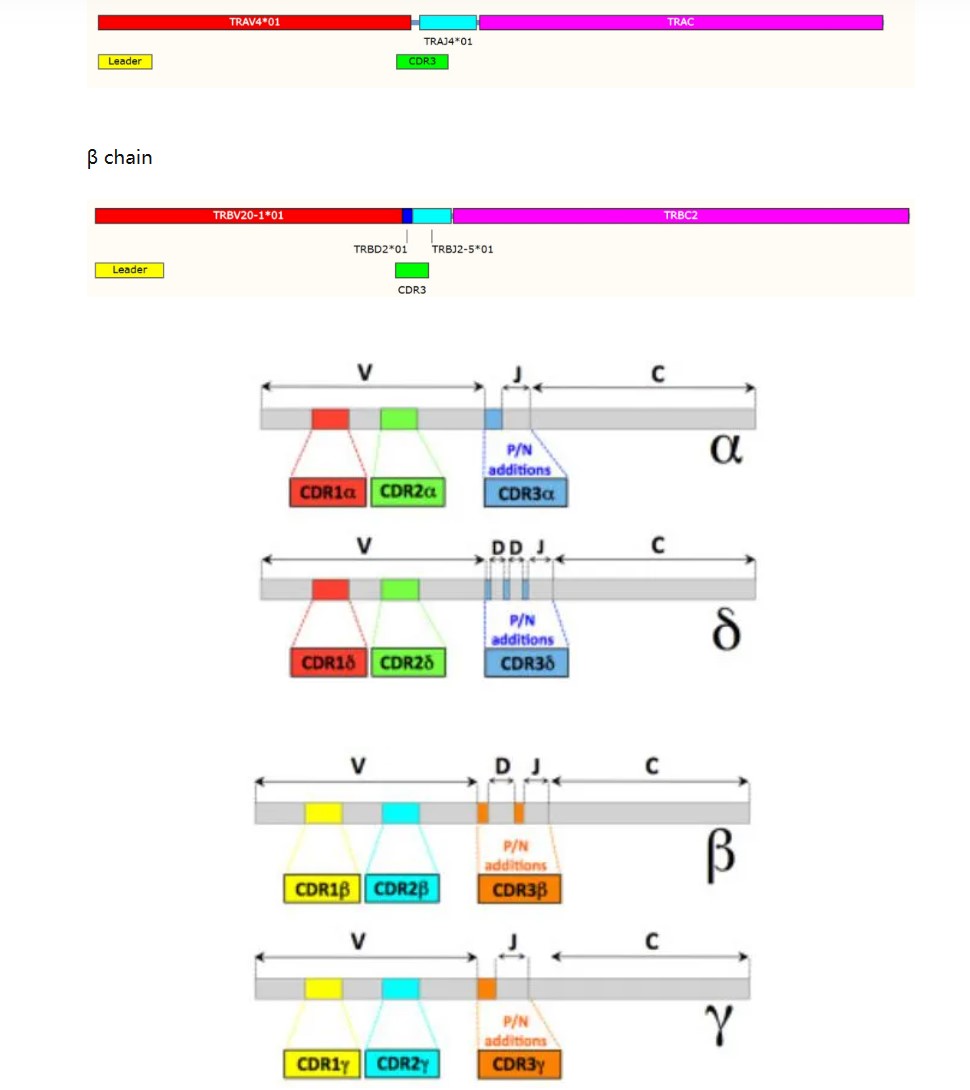
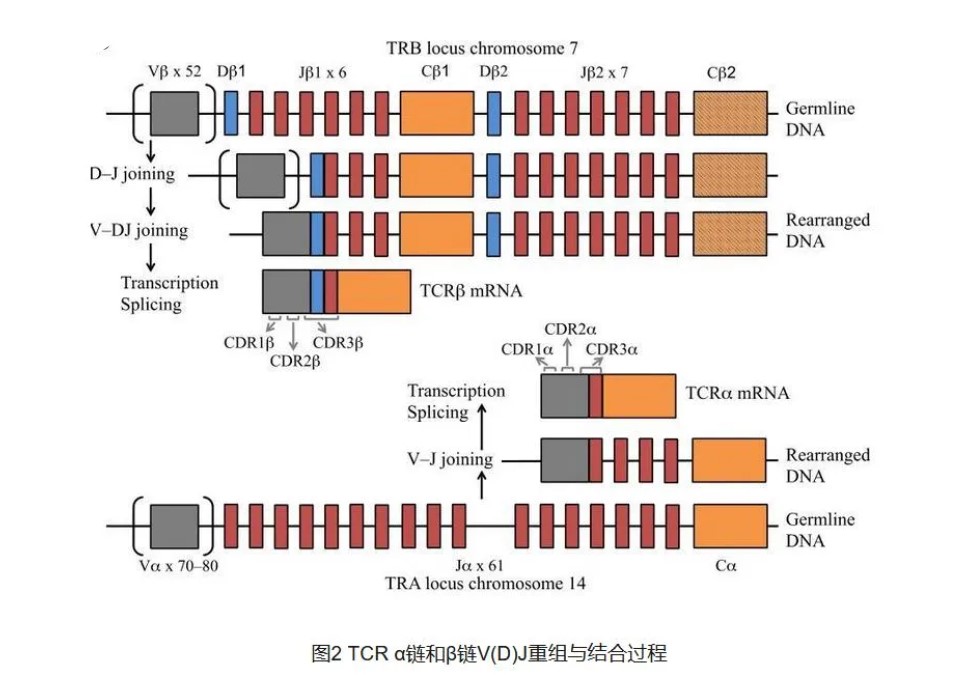
TCR 复合物的基本功能是引导 T 细胞识别特定的抗原。面对多种多样的抗原类型如何做到“特异”识别,这
就归功于 TCR 的“高度多态性”了。TCR
α链和γ链由一系列可变(variable,V)和连接(joining,J)基因编码,TCR
β和δ链还由高变(diversity,D) 基因编码。基于 RAG1 和 RAG2 重组酶的
V-(D)-J 重组过程中,V、D、J 中一个随机等位基因会与其他区段等位基因重组,形成功能多样化的片段。在 V-D、D-J 于 V-J
基因片段之间的连接点处会随机进行核 苷酸的添加、缺失。此外,在转录过程中位于重组 V(D)J 区域的序列以及恒定(Constant,
C)区域的基 因序列会进行可变剪切,形成存在一定功能差异的转录本。上述过程会导致超强的重组与结合多样 性,产生高可变型、高多样性的 T
细胞受体,确保对各类抗原进行鉴定。基于数学计算,理论上存在 10^12-10^15 种不同的 TCR[5],实际人体存在的 TCR
多样性大约在 10^13 这个数量级,说明看似随机的 TCR 重组并不是完全随机,可能受到某些机制的约束。产生 TCR 多样性的重组过程,是
T 淋巴细胞在初级淋巴器官——胸腺中的发育早期所特有。每个 TCR 链 都含有 3 个高度多样性的
loop,称为互补决定区域(complementarity determining re-gions, CDR1-3)。CDR1 和 CDR2
由 V 基因编码,而 CDR3 由 V-J 或 V-D-J 基因结合区域编码,高度多样化。此外,CDR3 是 TCR 直接接触抗原肽的区域,对
TCR 与抗原肽-MHC 复合体的互作起到决定性作用。正因为如 此,CDR3 是确定 T 细胞克隆类型的重要区域,两个 T
细胞不太可能表达相同的 CDR3 序列,除非它们来 自相同的 T 细胞克隆。
PART 2
CAR-T的每个元件的功能
抗原
(靶点)的选择一直是CAR-T领域的研究重点,如果仅仅有合适的抗原,缺乏共刺激信号,T细胞往往会未老先衰,起不到抗肿瘤的作用。在CAR-T的结构
中,任何一个部件都起着关键作用,下面我们就来看看CAR-T结构组成及其功能。
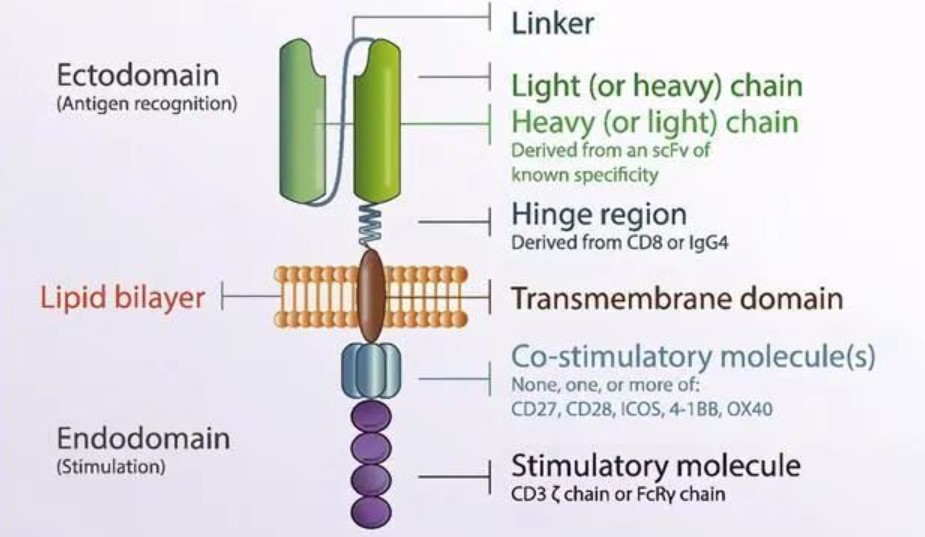
嵌合抗原受体 (CAR) 常规表达载体:
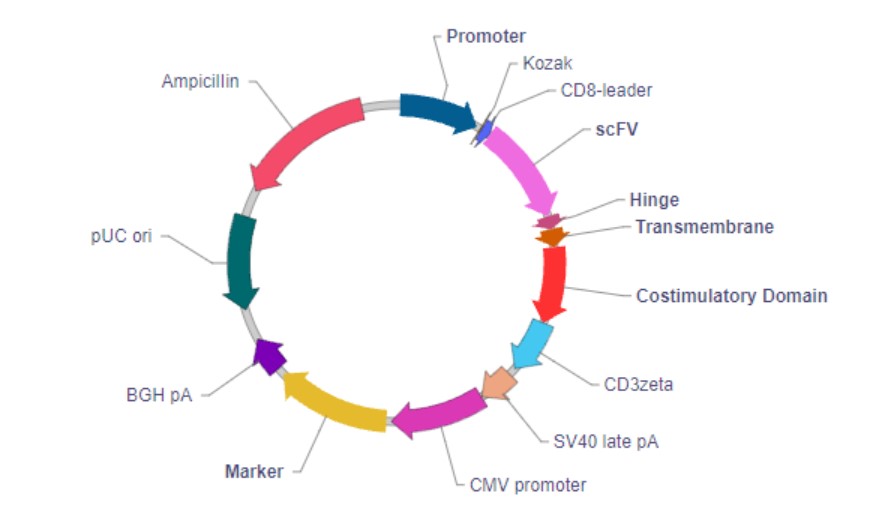
按照每个元件所处的位置,CAR主要分为三个功能区域,分别是:胞外结构域 、 跨膜结构域 和 胞内结构域 。
胞外结构域由负责识别并结合抗原的单克隆抗体的单链可变片段(single-chain variable
fragment,scFv)及一段起连接作用的铰链区(Hinge)构成。
胞内结构域由共刺激结构域(Costimulatory Domain)和信号转导结构域(Signaling Domain)构成。
PART
3
胞外结构域
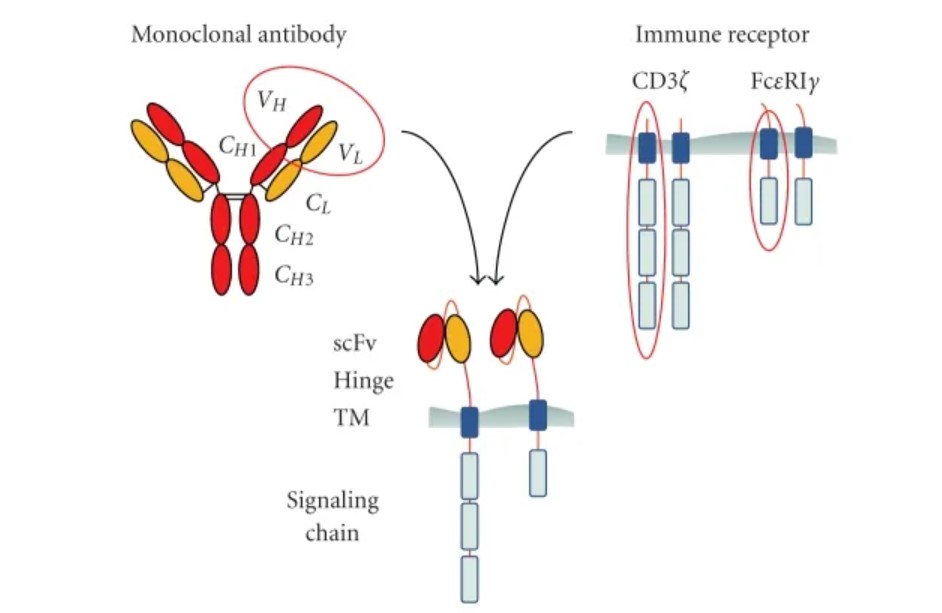
1. 抗原识别结构域(scFv)
结构组成:抗原识别结构域是CAR特异性结合肿瘤抗原的基础,其主要结构是scFv,
由单克隆抗体的轻链(VL)和重链(VH)通过多肽连接而成,保留有抗体对抗原的特异性和亲和力。
功能: scFv赋予了T细胞特异性识别并结合靶抗原的能力,与未经修饰的天然TCR-T细
胞相比,其对靶抗原的亲和力显著提高。CAR-T细胞通过scFv对靶抗原的识别结合不依赖MHC抗原呈递,一方面避免了肿瘤细胞通过调节MHC分子发生
逃逸,另一方面赋予了CAR-T细胞识别非肽抗原的能力。
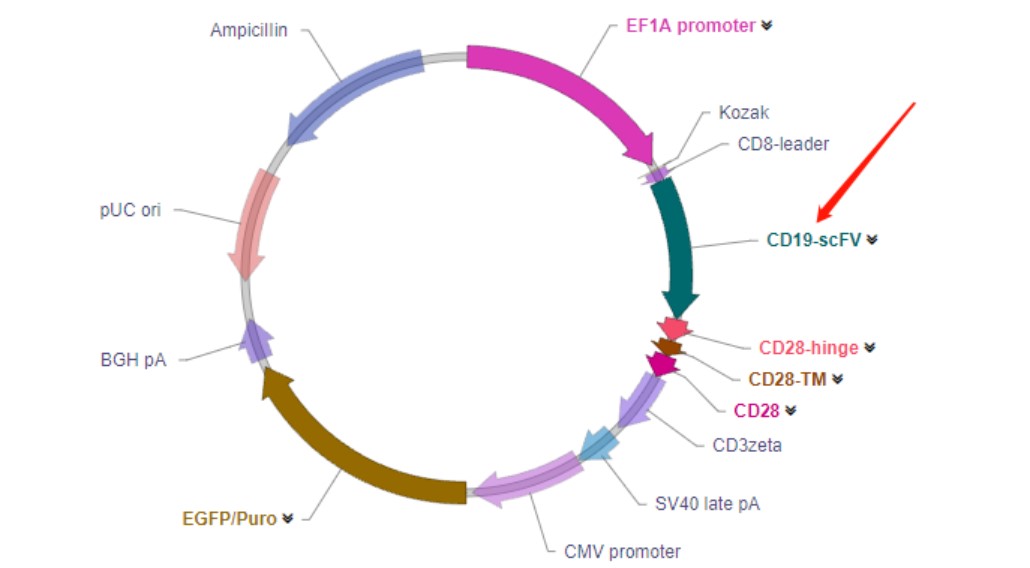
研究进展:目前大部分CAR-T研究是靶向TAA的,如包括CD19、CD20、CD22、
CD30、CD33、BCMA等,也就是说临床上主要是针对这些靶点来开发抗体。不过目前CAR-T临床研究所用的scFv多源于小鼠,这无形中增加了宿
主对CAR-T细胞的免疫排斥反应,发生细胞毒性或降低了CAR-T的持续性。因此,开发人源化或全人源的抗体来源的scFv,降低CAR-T细胞的免疫
原性,是CAR-T研究面临的关键问题之一。
2. 铰链区(Hinge)
结构组成: 铰链区连接scFv和跨膜结构域,大多数CAR的铰链区由IgG的铰链或
CD8α/CD28胞外区衍变而来。铰链区的长度取决于靶细胞抗原表位的位置及暴露程度。
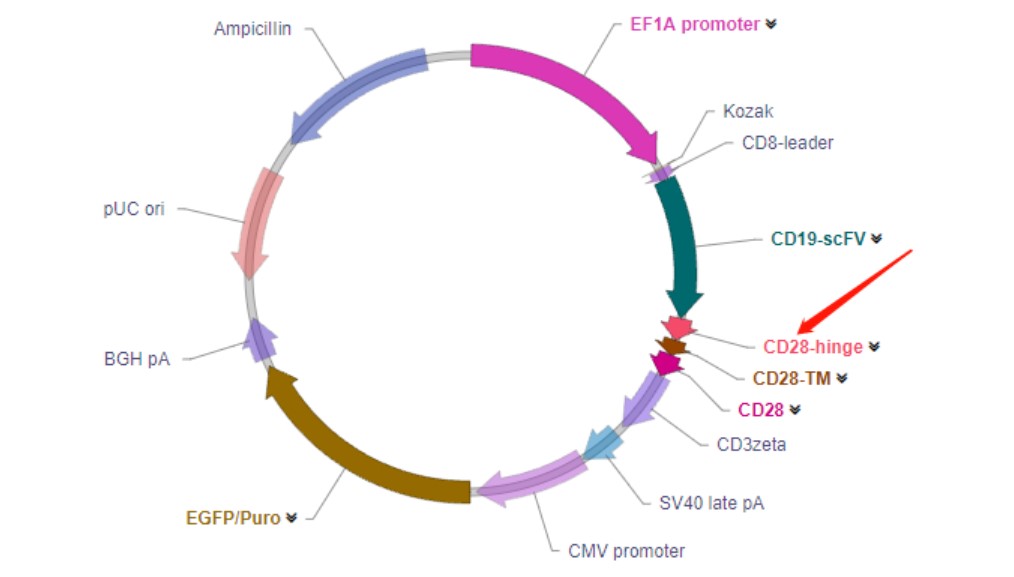
功能:多项研究表明,CAR-T细胞活化与铰链区长度有关。调整铰链区的长度可以使
CAR-T细胞和靶细胞处于最佳距离,在抗原抗体结合过程中避免大型磷酸酶的作用减弱CAR信号。然而,在某些情况下,抗原表位可能相对不可接近,需要使
用更长的铰链区,使scFv可以克服空间位阻,有效结合抗原。因此,抗原表位不同,铰链区的最佳长度也不同,在靶向新抗原时,可能需要相应地调整铰链区的
长度。
PART
4
跨膜结构域(Transmembrane region)
结构组成: 跨膜结构域将CAR的细胞外结构域与细胞内信号转导结构域连接,并将受体
锚定到T细胞膜上。常用的跨膜结构域来源于CD4,CD8α,CD28和CD3ζ。。
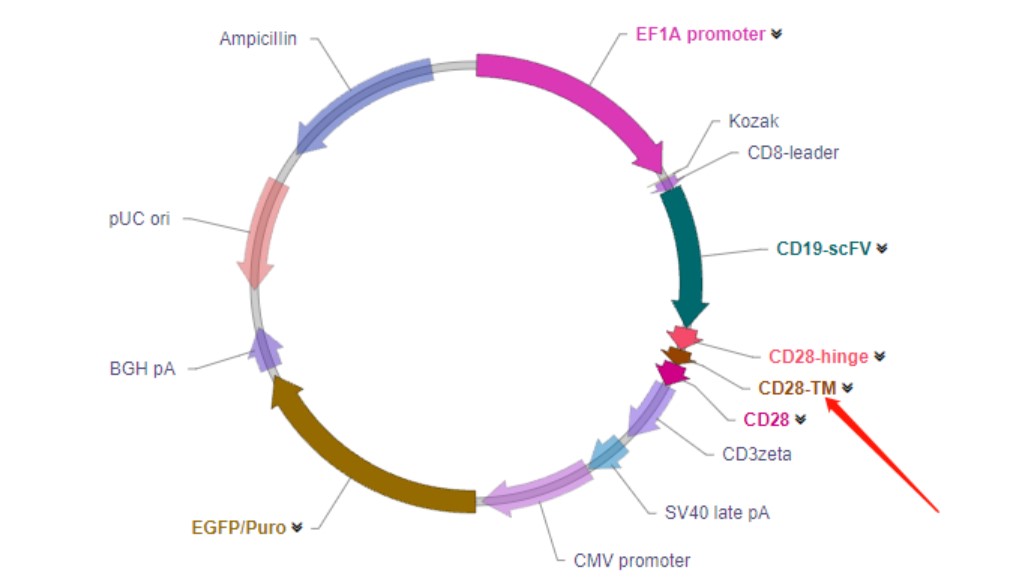
功能: 如CD3ζ跨膜域能够使CAR形成同源二聚体或与内源TCR形成异二聚体,增强
CAR-T细胞活性,但也正是由于其不需要与内源TCR结合即可高度活化T细胞,而逐渐被弃用。CD8α和CD28的跨膜域由于能够
促进CAR在细胞表面表达,目前在多数临床试验中采用。
PART 5
胞内结构域
1. 共刺激域(Costimulatory Domain)
结构组成: 共刺激域通常来自CD28受体家族(CD28,ICOS)或肿瘤坏死因子受体家族(4-1BB、OX40、CD27)。
功能:
共刺激域可实现协同刺激分子和细胞内信号的双重活化,使T细胞持续增殖并释放细胞因子,提高T细胞的抗肿瘤能力。如CD28共刺激域使CAR-T细胞依靠
糖酵解代谢,促使CAR-T细胞向效应T细胞分化。而4-1BB共刺激域促进线粒体生成,增强呼吸作用和脂肪酸氧化,抗原刺激后,CAR-T细胞优先分化
为中央记忆T细胞。

研究进展: 不过究竟是哪个共刺激结构域更好?科学家们仍在一直探索,如2018年,《Science
Signaling》期刊上一篇文章比较了这两种结构,结果发现:CD28表现出更快、更强的信号活动,而4-1BB则相对更慢、更温和。但是,4-
1BB有着“延长T细胞寿命并维持其抗癌作用”的潜在优势。
2. 信号转导结构域(Signaling Domain)
结构组成:
信号转导结构域通常为T细胞受体TCR/CD3ζ链或免疫球蛋白Fc受体FcεRIγ
链,含有免疫受体酪氨酸活化基序(immunoreceptor tyrosine-based activation motifs,ITAMs)。
功能: 发挥T细胞信号转导功能。
读完这篇文章的内容,对于CAR-T载体上CAR的每个元件的功能你是否都理解了呢?